
Hundur 3/2009
ISLANNINLAMMASKOIRIEN PERINNÖLLINEN MONIMUOTOISUUS
Teksti: Alina Niskanen
Tutkin pro gradu -työssäni vuonna 2008 Suomen islannin-lammaskoirien MHC-geenien monimuotoisuutta. Tutkimuksen taustaa ja tuloksia, sekä tarkempaa selostusta perinnöllisyys-tieteen termeistä, voi lukea Hundurista (2/2008) ja Koiramme-lehdestä (9/2008). Jatkan nyt väitöskirjassani muiden koira-rotujen ja suden lisäksi myös islanninlammaskoirien perinnöllisen monimuotoisuuden kartoitusta. Osa tekstistä on lainaa Koiramme-lehden (7-8/2009) jutustani, mutta lisänä on vertailua Islannissa muutama vuosi sitten tehtyyn monimuotoisuustutkimukseen. MHC-geenien tuotteet osallistuvat immuuni-puolustusreaktion alkuvaiheessa elimistölle vieraiden aineiden tunnistukseen. Tunnistettavia taudinaiheuttajia on lukuisia, joten MHC-geenien monimuotoisuus on eläimen terveyden kannalta erittäin tärkeää. Islanninlammaskoirilta ja muutamilta muilta roduilta tutkittiin kolmen vierekkäin sijaitsevan MHC-geenin monimuotoisuutta. Vaikka MHC-geeneillä on olennainen tehtävä immuunipuolustuksessa, niitä tutkimalla saadaan tietoa vain hyvin rajatun alueen monimuotoisuudesta. Jotta rodun perinnöllisen monimuotoisuuden tilasta olisi saatu laajempaa tietoa, tutkittiin islanninlammaskoirilta myös niin kutsuttuja mikrosatelliittimarkkereita eri puolilta perimää. Mikrosatelliitit ovat DNA:n toistojaksoja, jotka koostuvat usein kahden emäksen toistoista, esimerkiksi CACACA. Saman mikrosatelliitin eri alleelit eroavat toisistaan toistojen määrän suhteen, eli alleelit ovat eripituisia. Mikrosatelliitit eivät koodaa ainuttakaan geenituotetta, eikä niillä siis ole yhteyksiä immuunipuolustukseen. Ne ovat eliön kannalta todennäköisesti melko turhia ja syntyvät, muuttuvat ja säilyvät populaatiossa sattuman kautta. Päinvastoin kuin MHC-geeneihin, luonnon-valinnan ei oleteta vaikuttavan mikrosatelliittien monimuotoisuuteen. Niiden monimuotoisuuteen vaikuttavat pääasiassa vain populaatiokoon muutokset tai jalostustottumukset, kuten sukusiitoksen määrä. Tästä johtuen mikrosatelliitteja käytetään paljon eri lajien ja myös koirarotujen perinnöllisen muuntelun tutkimiseen. Lisäksi niitä käytetään muun muassa isyys-testaukseen, koska jokaisen yksilön mikrosatelliiteista muodostuva geneettinen sormenjälki on erilainen.
Tulokset mikrosatelliittimarkkereilla
Suomen islanninlammaskoirapopulaation perinnöllinen monimuotoisuus on matalampi kuin koirarotujen keskimäärin (taulukko 1). Tulos on yllättävä, sillä Islannissa muutama vuosi sitten tehdyssä tutkimuksessa rodun monimuotoisuuden taso havaittiin melko korkeaksi. Heterotsygoottinen tarkoittaa eriperintäistä, eli yksilö on saanut erilaisen geenin muodon (alleelin) kummaltakin vanhemmaltaan. Jos rodussa esiintyvät alleelit jakaantuvat yksilöihin ideaalipopulaation tavoin, populaation heterotsygotia-aste noudattaa niin kutsuttua odotettua heterotsygotiaa (He). Odotettua heterotsygotia-astetta laskettaessa lähtöarvoina käytetään alleelien lukumäärää ja yleisyyttä. Odotettu heterotsygotia-aste kertoo teoreettisesta populaation tasapainotilanteesta, joten sitä voidaan käyttää eri populaatioiden väliseen vertailuun, jos halutaan olla huomioimatta esimerkiksi sukusiitoksen vaikutuksia. Selkeästi suku-siitoksesta kärsivissä roduissa odotettu heterotsygotia-aste kertoo heterotsygotian tason, joka rodussa olisi mahdollista saavuttaa järkevällä jalostuskäytöllä.
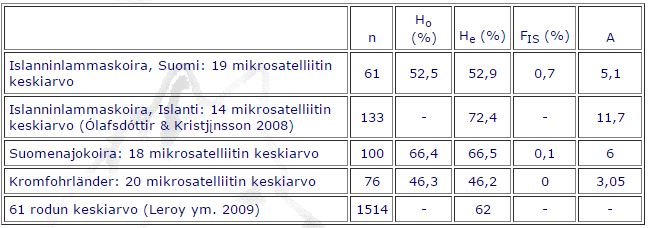
Islanninlammaskoirien Suomen populaation odotettu heterotsygotia oli vain 52,9 %, kun taas Islannin populaation odotettu heterotsygotia oli 72,4 %. Keskimääräinen alleelien lukumäärä oli Suomen populaatiossa 5,1 ja Islannin populaatiossa 11,7. Havaittujen alleelien määrään vaikuttaa tutkittujen yksilöiden määrä, joten arvot eivät ole täysin vertailukelpoisia erikokoisilla näytemäärillä. Tutkimukset tehtiin lähes täysin eri mikrosatelliittimarkkereilla, mutta silti tulokset poikkeavat toisistaan yllättävän paljon. Taulukossa 1 on esitetty tulokset myös suomenajokoirien ja kromfohrländerien monimuotoisuus-tutkimuksista. Suomenajokoiralla oli sekä runsaampi alleelimäärä (6) että korkeampi heterotsygotia-aste (66,5 %) kuin islanninlammaskoiralla. Sen sijaan kromfohrländer oli perimältään vähemmän monimuotoinen; sen odotettu heterotsygotia-aste oli vain 46,2 % ja alleelien lukumäärä 3,05. Islanninlammaskoiran ja kromfohrländerin odotettu heterotsygotia-aste on matala myös verrattuna 61 rodun keskiarvoon (62 %).
Sukusiitos ja sukusiitoskerroin ovat käsitteinä monille koiraharrastajille hyvinkin tuttuja. Oman koiransa sukutaulun perusteella lasketun sukusiitoskertoimen näkee esimerkiksi Suomen Kennelliiton Koiranetistä. Geneettisen aineiston avulla popu-laation sukusiitoskerroin (FIS) voidaan laskea vertaamalla havaitun ja odotetun heterotsygotian määrää toisiinsa. Positiivinen sukusiitoskerroin kertoo homotsygoottien ylimäärästä, eli sukusiitoksesta, ja negatiivinen arvo homotsygoottien alimäärästä, eli “ulkosiitoksesta”. Tulosten perusteella sen enempää islanninlammaskoira (FIS = 0,7 %) kuin muutkaan tutkitut rodut eivät ole erityisen sukusiittoisia. Kaikki islannin-lammaskoirat ovat sukua toisilleen, joten sukusiitokselta ei voi välttyä. Vaikuttaisi kuitenkin siltä, että jalostukseen käytetyt koirat eivät ole keskenään läheisempää sukua kuin koko tutkittu kanta.
Taulukko 1.
Islanninlammaskoiran Suomen ja Islannin populaation, suomenajokoiran ja kromfohrländerin mikrosatelliittimarkkerien avulla havaittu heterotsygotia-aste (Ho), tasapainotilanteessa odotettu heterotsygotia-aste (He), sukusiitosaste (FIS), alleelien lukumäärä (A) ja tutkittujen koirien määrä (n).
Kertausta MHC-tuloksista
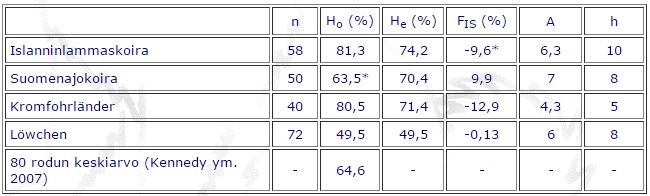
Vertailun vuoksi taulukossa 2 on jo aiemmin julkaistut MHC-geenien monimuotoisuustulokset. Islannin populaatiosta ei ole tehty MHC-tutkimusta. Suomenajokoiraa lukuun ottamatta kaikilla tutkituilla roduilla on negatiivinen sukusiitoskerroin, tosin tulos on tilastollisesti merkitsevä vain islanninlammaskoirilla (-9,6 %). Negatiivinen sukusiitoskerroin kuvaa “ulkosiitosta”, eli populaatioon tulee lisääntyviä yksilöitä toisesta populaatiosta. Tulos vaikuttaisi Suomen kannassa varsin järkevältä, koska ulkomaantuonteja on paljon jalostuskäytössä. Kuitenkaan mikrosatelliiteilla tutkittuna ei havaita negatiivista sukusiitoskerrointa, joten selitys on hieman ontuva. Islanninlammaskoira on MHC-geeneillä mitattuna tutkituista roduista monimuotoisin. Rodulla on korkein havaittu ja odotettu heterotsygotia-aste sekä alleelien ja haplotyyppien lukumäärä.
Taulukko 2.
DLA-DRB1-, DQA1- ja DQB1-geenien muuntelun mittojen keskiarvot islanninlammaskoirilla, suomenajokoirilla, kromfohrländereillä ja löwcheneillä. Tutkittujen koirien määrä (n), havaittu heterotsygotia-aste (Ho), tasapainotilanteessa odotettu heterotsygotia-aste (He), sukusiitosaste (FIS), alleelien lukumäärä (A) ja haplotyyppien lukumäärä (h). (Tilastollisesti merkitsevä poikkeama: * p < 0,05)
Pohdinta
Ero Islannin ja Suomen islanninkoirapopulaatioiden heterotsygotiassa oli yllättävä. Islannissa jalostuskäytössä on hyvin pieni osa koirista ja urosten matadorikäyttö on yleistä. Suomessa sen sijaan pyritään uroksia ja narttuja käyttämään tasaisesti jalostukseen. Pentueen vanhemmat valitaan useimmiten myös mahdollisuuksien mukaan erisukuisista koirista. Päinvastoin kuin voisi olettaa, odotettu heterotsygotia-aste oli huomattavasti matalampi Suomessa kuin Islannissa. Siksi tuntuu vaikealta uskoa, että nyt saadut tulokset kertoisivat todellisen tilanteen. Toisaalta tiedossani ei tällä hetkellä ole Islannin populaation havaitun heterotsygotian keskiarvoa. Se kertoisi nimenomaan populaation viimeaikaisista tapahtumista. Olisi erittäin mielenkiintoista verrata populaatioita samoilla mikrosatelliittimarkkereilla, jolloin voitaisiin sulkea pois mahdolliset markkereista johtuvat erot. Nyt tutkimuksissa oli käytössä vain kaksi samaa mikrosatelliittia, joiden odotettu heterotsygotia-aste oli korkeampi Suomessa kuin Islannissa (tulokset eivät ole näkyvillä).
Aiemman oletuksen mukaan islanninlammaskoirien MHC-geenien korkea heterotsygotia-aste johtuisi tuontikoirien runsaasta jalostuskäytöstä. Mikrosatelliiteilla mitattuna heterotsygotia-aste oli kuitenkin pikemminkin odotettua matalampi. Islanninlammaskoira ei myöskään ollut heterotsygoottisempi kuin muut tutkitut rodut. Syytä näin suurelle erolle eri tutkimusmuotojen välillä on vaikea sanoa. Yksi mahdollinen selitys voisi olla nimenomaan MHC-geeneihin kohdistuva luonnonvalinta. Toisaalta näin voimak-kaan valinnan pitäisi näkyä jossain määrin myös mikro-satelliiteilla tutkittuna. Myös kromfohrländerit olivat monimuotoisempia DLA-geeneillä kuin mikrosatelliiteilla mitattuna. Vastaavaa on havaittu aiemmin luonnoneläimillä, esimerkiksi kettujen saaripopulaatiossa. Kettupopulaatio oli luultavasti kutistunut muutamaan yksilöön ja vaikka populaatiokoko oli tutkimushetkellä jo kasvanut, ei mikrosatelliiteilla havaittu lainkaan heterotsygotiaa. MHC-geeneissä havaittiin kuitenkin huomattavaa heterotsygotiaa. Selityksenä pidetään MHC-geeneihin kohdistuvaa luonnonvalintaa, joka säilyttää heterotsygotiaa, vaikka sitä ei esiinny valinnan suhteen neutraaleissa mikrosatelliiteissa. Myös koirien kohdalla mahdollinen selitys MHC-geenien monimuotoisuudelle on heterotsygotiaa ylläpitävä valinta. Luonnoneläimiin verrattuna lemmikkikoirien tilanne on kimurantimpi, sillä koirat ovat ihmisen hoidossa syntymästään alkaen ja kaiken lisäksi ihminen valitsee koiran pariutumiskumppanin. Käytännössä koira on luonnon armoilla vain astutuksen ja syntymän välisenä aikana. Tämä tulos herättää mielenkiintoisia kysymyksiä, joihin etsitään vastauksia tutkimuksen tulevissa vaiheissa.
Lähteitä
Aguilar, A., Roemer, G., Debenham, S., Binns, M., Garcelon, D. & Wayne, R. K. 2004: High MHC diversity maintained by balancing selection in an otherwise genetically monomorphic mammal. – Proceedings of the National Academy of Sciences USA. 101: 3490-3494.
Kennedy, L. J., Barnes, A., Short, A., Brown, J. J., Lester, S., Seddon, J., Fleeman, L., Francino, O., Brkljacic, M., Knyazev, S., Happ, G. M. & Ollier, W. E. R. 2007: Canine DLA diversity: 1. New alleles and haplotypes. – Tissue antigens. 69: 272-288.
Leroy, G., Verrier, E., Meriaux, J. C. & Rognon, X. 2009: Genetic diversity of dog breeds: within-
breed diversity comparing genealogical and molecular data. – Animal Genetics. 40: 323–332.
Niskanen, A. 2008. MHC II -luokan geenien monimuotoisuus Suomen susipopulaatiossa (Canis
lupus) ja islanninlammaskoirapopulaatiossa (Canis familiaris). Pro gradu -tutkielma. Oulun yliopisto, biologian laitos, 69 s.
Ólafsdóttir, G. A. & Kristjįnsson, T. 2008. Correlated pedigree and molecular estimates of inbreeding and their ability to detect inbreeding depbression in the Icelandic sheepdog, a recently bottlenecked population of domestic dogs. – Conservation genetics. 9: 1639-1641.